SCFAs Part 2: The benefits of butyrate
Did you know that butyrate, a metabolite produced in your gut, has benefits for the brain, skin, immune system, and more?

In the last installment of the short-chain fatty acid (SCFA) series, I covered the basics of SCFAs, including how they are produced, where they traffic in the body, and their varied functions. In this second article, I’ll focus on the 4-carbon SCFA butyrate, and discuss the many benefits it has on host health.
Butyrate protects the gut
Butyrate is the preferred substrate for colonocytes, providing 60-70% of the energy requirements for colonic epithelial cells1,2. Butyrate suppresses colonic inflammation,3 is immunoregulatory in the gut,4 and improves gut barrier permeability by accelerating assembly of tight junction proteins.5,6
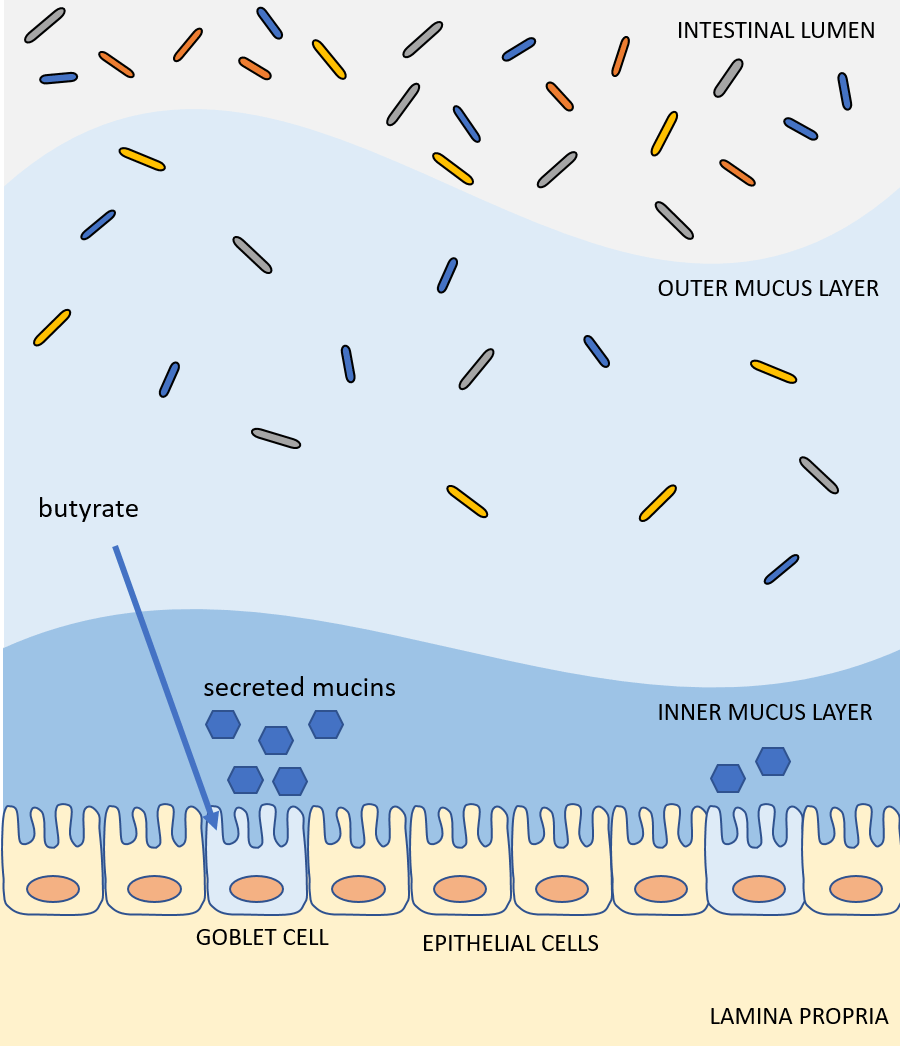
Butyrate also influences the mucus layer. A healthy colonic epithelium is coated in a double layer of mucus. The thick, inner layer is dense and largely devoid of microbes, protecting the epithelium from contact with commensals and pathogens alike. The loose, outer layer of mucus is home to many bacteria, some of which feed on the glycoproteins of the outer mucus layer itself. Both of these mucus layers are organized by the MUC2 mucin protein, which is secreted by goblet cells in the epithelium. Supplementation of physiological concentrations of butyrate has been shown to increase MUC2 gene expression and MUC2 secretion in a human goblet cell line.7,8
Butyrate improves metabolic function
Butyrate also influences metabolism. In a mouse model of obesity, supplemental butyrate was shown to improve insulin sensitivity, increase energy expenditure, and reduce adiposity. It also increased the number of mitochondria and their activity in skeletal muscle and brown fat tissue.9 This is not surprising, as butyrate responsive receptors are highly expressed in adipose tissue. This expression declines with age, and may explain the age-associated decline in insulin sensitivity.10,11 Butyrate also induces the production of satiety hormones, which reduce food intake.12
Butyrate may also have benefits for other metabolic diseases. In rats, oral butyrate given to young mice early in life showed a trend towards delaying the development of diabetes13. Butyrate supplementation has also been shown to attenuate diet-induced fatty liver disease in mice by improving gut barrier function and correcting microbial dysbiosis14.
Butyrate alters gene expression
While our genetic makeup is essentially unchangeable, the epigenetics, or expression of our genes, is quite malleable. In the nucleus of every cell in our body, DNA is wound around histone proteins. When the histones of a gene are deacetylated, the histones and their associated DNA tend to pack very tightly. This makes it very difficult for any enzymes to bind to transcribe the message encoded in the DNA. The gene will not be made into a protein, and is functionally “off”. On the other hand, when the histones of a gene are acetylated, the histones and their associated DNA tend to pack more loosely. This allows enzymes to bind to the promoter region of the gene and initiate its transcription. This is the first step towards making a functional protein from the gene.
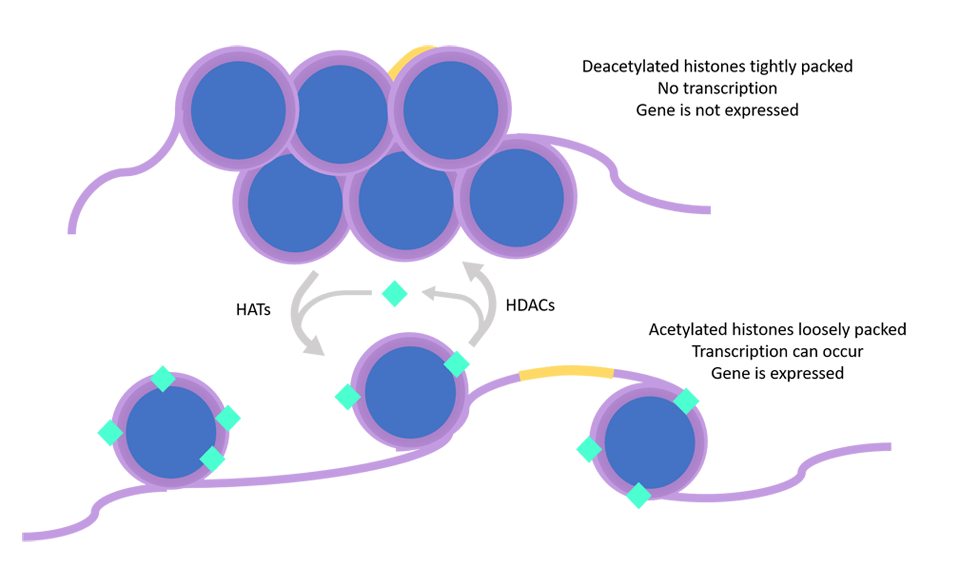
So, what determines if a gene is acetylated or deacetylated? Enzymes called histone acetyltransferases (HATs) add acetyl groups to histones, while enzymes called histone deacetylases (HDACs) remove acetyl groups. Many signaling molecules in the body can influence the activity of these enzymes. One such molecule is…you guessed it…butyrate! Butyrate increases acetylation in two different ways, for distinct sets of genes. First, butyrate is considered a potent HDAC activity inhibitor, meaning that it prevents HDACs from removing acetyl groups (Boffa 1978, Hinnebusch, 2002, Kiefer 2006, Waldecker 2008). For other genes, butyrate enhances HAT activity. Both of these mechanisms serve to increase acetylation and keep genes turned “on”. In the next section, we’ll see how this is put into action in cancer cells.
Butyrate is protective against cancer
Butyrate has long been known to stimulate epithelial cell proliferation, yet at the same time, be protective against colon cancer. In other words, butyrate could exert different effects in healthy and cancerous colonic cells.
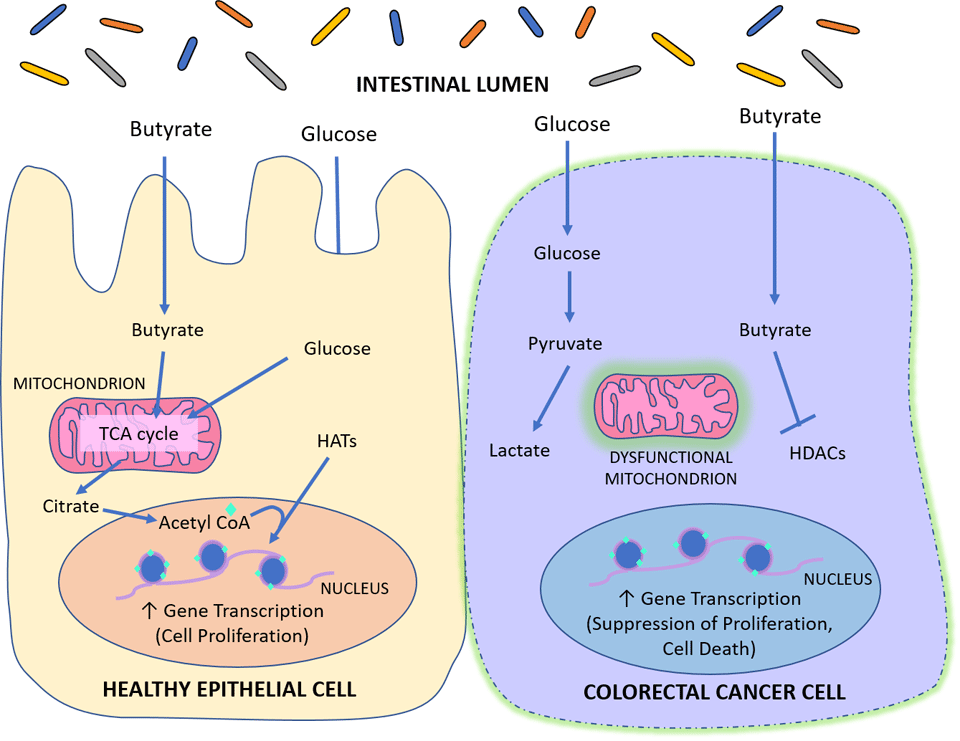
Only recently was the mechanism behind this discovered. In an elegant set of experiments, it was determined that the Warburg effect could explain the paradoxical nature of butyrate. The Warburg effect is a phenomenon that involves the metabolism of rapidly-dividing cancerous cells. In a normal epithelial cell, fatty acids like butyrate are largely metabolized via the tricarboxylic acid (TCA) cycle in the mitochondria for energy. An increase in flux through the TCA cycle results in the production of citrate in the cell cytosol, which can be converted to acetyl CoA in the nucleus. This acetyl CoA provides the acetyl group necessary for HATs to acetylate the histones of genes that promote epithelial cell proliferation.
Cancerous cells, on the other hand, tend to have dysfunctional mitochondria, meaning that they can’t metabolize fatty acids (like butyrate) for energy. Because of this, they rely largely on glucose metabolism for their energy production and fatty acids like butyrate tend to accumulate in the cell. This increased cellular butyrate concentration results in increased HDAC inhibition, and the expression of genes that prevents cancer cells from replicating.15
This phenomenon isn’t unique to colon cancer. Butyrate has been identified as having anticancer activity in a variety of human cancer cell lines, including tongue cancer,16 prostate cancer,17,18 liver cancer,19,20 breast cancer,21 lung cancer,22,23 and neuroblastoma.24
Butyrate may benefit the brain
Unfortunately, most of our butyrate-and-the-brain research is limited to animal models. Still, butyrate has been shown to have widespread and dramatic effects on brain health. Mice given sodium butyrate or the butyrate-producing bacteria Clostridium butyricum showed increased neurogenesis, reduced oxidative stress, and improved recovery following ischemic brain injury.25,26 Sodium butyrate has also been shown to restore the integrity of the blood-brain barrier and attenuate neurological deficits following traumatic brain injury in mice.27 Additionally, butyrate may prevent nerve degeneration in the autosomal recessive disease spinal muscular atrophy.28
Butyrate has shown anti-depressant activity in mice, altering expression of genes like brain-derived neurotrophic factor (BDNF) in the hippocampus and frontal cortex and increasing serotonin levels.29,30 It also reduces manic-like behavior in a drug-induced model of bipolar disorder31, and increases expression of neurotrophic factors and reduces cognitive impairment in models of chronic mild stress and maternal separation.32
Butyrate also has interesting implications for memory formation and retention. Disturbances in hippocampal gastrin-releasing peptide receptors (GRPRs) may inhibit proper memory formation and extinction and contribute to the cognitive impairment associated with neurodevelopmental disorders. One study in mice found that HDAC inhibition by butyrate could attenuate the cognitive impairment that results from GRPR signaling blockade.33 Another study found that, like exercise, butyrate places the brain in a “state of readiness for plasticity”.34
Lastly, butyrate may have benefits for autism spectrum disorder. I’ll be discussing this more in part 5 when I discuss SCFA imbalance and the propionate-ASD connection.
Butyrate regulates the immune system
Butyrate also plays a major role in regulating the immune system, and butyrate responsive receptors have been found on virtually every type of immune cell.35
Butyrate signaling results in an overall anti-inflammatory immune profile, inhibiting production of proinflammatory cytokine and upregulating production of anti-inflammatory cytokines36,37. It has been shown to do this even in the face of an inflammatory insult, such as lipopolysaccharide (LPS) from the cell walls of gram-negative bacteria. This may occur through HDAC inhibition of the NF-kB pathway, a major pathway involved in the release of inflammatory cytokines38,39. In fact, butyrate mediated HDAC inhibition has been observed in many different immune cell types.40
Butyrate also promotes the generation of colonic and peripheral regulatory T cells (Tregs), which serve to help dampen the immune response,41–43 influences both Treg and effector T cell function,44,45 and induces activity of cathelicidin, a potent antimicrobial known to play an important role in innate defense against bacterial infection, in colonic and lung epithelial cells.46,47
Butyrate and the skin
Of particular interest to me, due to my previous history with eczema and laboratory research on the gut-skin axis, is the effect of butyrate on skin health. Butyrate acts as a regulator of the skin immune system. Subcutaneously or topically applied butyrate was shown to reduce contact hypersensitivity, likely explained by the observed increase in regulatory T cells and reduction in inflammatory effector T cells.48 Butyrate also induces the synthesis of collagen, an important structural component of skin.49
Butyrate and bone health
Butyrate may also improve bone health. Bone is constantly being remodeled in a delicate balance between osteoblast (bone-building) cell activity and osteoclast (bone breakdown) cell activity. Mesenchymal stem cells (MSCs), which reside in the bone marrow, can differentiate into osteoblasts (bone-building cells) or adipocytes (fat cells). One study found that sodium butyrate caused MSCs to preferentially differentiate into osteoblasts, suggesting that it could promote bone formation.50 Another study found that osteoblasts cultured with butyrate had increased number and calcium content of mineralized nodules. They also expressed higher levels of osteoprotegerin, a protein that serves to reduce osteoclast (bone breakdown) activity. 51 In animal models, a mixture of SCFAs that includes butyrate has been shown to increase circulating levels of insulin-like growth factor (IGF) and promote bone formation.52
Ways to increase butyrate
To sum up, we’ve learned that butyrate is a potent modifier of gene expression, and has widespread benefits for host health. There are many reasons we might want to increase butyrate production, and there are several ways to do so:
- Increase the abundance of butyrate-producing bacteria. Bacterial species with particularly high capacities for butyrate production belong to both the Firmicutes and Bacteroidetes phyla and include species such as Clostridium leptum, Eubacterium spp., Roseburia spp., Coprococcus spp., and Faecalibacterium prausnitzii.53 Genera like Bifidobacteria and Lactobaccilus may also contribute to butyrate production by producing acetate and lactate, which other bacteria can then convert to butyrate.
- Increase the substrates available to butyrate-producing bacteria. This means eating prebiotic foods with fermentable fibers that stimulate butyrate production. Some of the most potently butyrogenic fibers include resistant starch and fructooligosaccharides. I’ll be discussing this more in SCFAs part 7: the definitive guide to fiber.
- Supplementation. This should be done with extreme caution, and with the right supplement form. Which brings me to my final point for today…
Is butyrate always good?
Nope! This is only part 2! In the next article of this series, I’ll talk about how excess butyrate can be toxic, particularly in conditions involving gut inflammation. I’ll also discuss the various forms of supplementation, and why most butyrate supplements may do more harm than good. Be sure to subscribe below!
Sources:
- Donohoe, D. R. et al. The Microbiome and Butyrate Regulate Energy Metabolism and Autophagy in the Mammalian Colon. Cell Metab. 13, 517–526 (2011).
- Suzuki, T., Yoshida, S. & Hara, H. Physiological concentrations of short-chain fatty acids immediately suppress colonic epithelial permeability. Br. J. Nutr. 100, 297–305 (2008).
- Singh, N. et al. Activation of Gpr109a, receptor for niacin and the commensal metabolite butyrate, suppresses colonic inflammation and carcinogenesis. Immunity 40, 128–139 (2014).
- Chang, P. V., Hao, L., Offermanns, S. & Medzhitov, R. The microbial metabolite butyrate regulates intestinal macrophage function via histone deacetylase inhibition. Proc. Natl. Acad. Sci. U. S. A. 111, 2247–2252 (2014).
- Peng, L., He, Z., Chen, W., Holzman, I. R. & Lin, J. Effects of butyrate on intestinal barrier function in a Caco-2 cell monolayer model of intestinal barrier. Pediatr. Res. 61, 37–41 (2007).
- Peng, L., Li, Z.-R., Green, R. S., Holzman, I. R. & Lin, J. Butyrate Enhances the Intestinal Barrier by Facilitating Tight Junction Assembly via Activation of AMP-Activated Protein Kinase in Caco-2 Cell Monolayers. J. Nutr. 139, 1619–1625 (2009).
- Burger-van Paassen, N. et al. The regulation of intestinal mucin MUC2 expression by short-chain fatty acids: implications for epithelial protection. Biochem. J. 420, 211–219 (2009).
- Hatayama, H., Iwashita, J., Kuwajima, A. & Abe, T. The short chain fatty acid, butyrate, stimulates MUC2 mucin production in the human colon cancer cell line, LS174T. Biochem. Biophys. Res. Commun. 356, 599–603 (2007).
- Gao, Z. et al. Butyrate improves insulin sensitivity and increases energy expenditure in mice. Diabetes 58, 1509–1517 (2009).
- Thangaraju, M. et al. GPR109A is a G-protein-coupled receptor for the bacterial fermentation product butyrate and functions as a tumor suppressor in colon. Cancer Res. 69, 2826–2832 (2009).
- Fink, R. I., Kolterman, O. G., Griffin, J. & Olefsky, J. M. Mechanisms of Insulin Resistance in Aging. J. Clin. Invest. 71, 1523–1535 (1983).
- Lin, H. V. et al. Butyrate and Propionate Protect against Diet-Induced Obesity and Regulate Gut Hormones via Free Fatty Acid Receptor 3-Independent Mechanisms. PLOS ONE 7, e35240 (2012).
- Li, N. et al. Butyrate and type 1 diabetes mellitus: can we fix the intestinal leak? J. Pediatr. Gastroenterol. Nutr. 51, 414–417 (2010).
- Zhou, D. et al. Sodium butyrate attenuates high-fat diet-induced steatohepatitis in mice by improving gut microbiota and gastrointestinal barrier. World J. Gastroenterol. 23, 60–75 (2017).
- Donohoe, D. R. et al. The Warburg effect dictates the mechanism of butyrate-mediated histone acetylation and cell proliferation. Mol. Cell 48, 612–626 (2012).
- Jeng, J.-H. et al. Toxic and metabolic effect of sodium butyrate on SAS tongue cancer cells: role of cell cycle deregulation and redox changes. Toxicology 223, 235–247 (2006).
- Kim, J., Park, H., Im, J. Y., Choi, W. S. & Kim, H. S. Sodium butyrate regulates androgen receptor expression and cell cycle arrest in human prostate cancer cells. Anticancer Res. 27, 3285–3292 (2007).
- Kuefer, R. et al. Sodium butyrate and tributyrin induce in vivo growth inhibition and apoptosis in human prostate cancer. Br. J. Cancer 90, 535–541 (2004).
- Nakamura, M. et al. Reduction of telomerase activity in human liver cancer cells by a histone deacetylase inhibitor. J. Cell. Physiol. 187, 392–401 (2001).
- Wakabayashi, K. et al. Gene expression associated with the decrease in malignant phenotype of human liver cancer cells following stimulation with a histone deacetylase inhibitor. Int. J. Oncol. 26, 233–239 (2005).
- M, M. & R, K. Bcl-2 expression regulates sodium butyrate-induced apoptosis in human MCF-7 breast cancer cells. Cell Growth Differ. Mol. Biol. J. Am. Assoc. Cancer Res. 7, 311–318 (1996).
- Pellizzaro, C., Coradini, D., Daniotti, A., Abolafio, G. & Daidone, M. G. Modulation of cell cycle-related protein expression by sodium butyrate in human non-small cell lung cancer cell lines. Int. J. Cancer 91, 654–657 (2001).
- Denlinger, C. E., Keller, M. D., Mayo, M. W., Broad, R. M. & Jones, D. R. Combined proteasome and histone deacetylase inhibition in non–small cell lung cancer. J. Thorac. Cardiovasc. Surg. 127, 1078–1086 (2004).
- Schneider, F. H. Effects of sodium butyrate on mouse neuroblastoma cells in culture. Biochem. Pharmacol. 25, 2309–2317 (1976).
- Kim, H. J., Leeds, P. & Chuang, D.-M. The HDAC inhibitor, sodium butyrate, stimulates neurogenesis in the ischemic brain. J. Neurochem. 110, 1226–1240 (2009).
- Sun, J. et al. Clostridium butyricum pretreatment attenuates cerebral ischemia/reperfusion injury in mice via anti-oxidation and anti-apoptosis. Neurosci. Lett. 613, 30–35 (2016).
- Li, H. et al. Sodium butyrate exerts neuroprotective effects by restoring the blood-brain barrier in traumatic brain injury mice. Brain Res. 1642, 70–78 (2016).
- Chang, J.-G. et al. Treatment of spinal muscular atrophy by sodium butyrate. Proc. Natl. Acad. Sci. U. S. A. 98, 9808–9813 (2001).
- Schroeder, F. A., Lin, C. L., Crusio, W. E. & Akbarian, S. Antidepressant-like effects of the histone deacetylase inhibitor, sodium butyrate, in the mouse. Biol. Psychiatry 62, 55–64 (2007).
- Sun, J. et al. Antidepressant-like effects of sodium butyrate and its possible mechanisms of action in mice exposed to chronic unpredictable mild stress. Neurosci. Lett. 618, 159–166 (2016).
- Varela, R. B. et al. Sodium butyrate and mood stabilizers block ouabain-induced hyperlocomotion and increase BDNF, NGF and GDNF levels in brain of Wistar rats. J. Psychiatr. Res. 61, 114–121 (2015).
- Valvassori, S. S. et al. Sodium butyrate functions as an antidepressant and improves cognition with enhanced neurotrophic expression in models of maternal deprivation and chronic mild stress. Curr. Neurovasc. Res. 11, 359–366 (2014).
- Petry, F. S. et al. Histone deacetylase inhibition prevents the impairing effects of hippocampal gastrin-releasing peptide receptor antagonism on memory consolidation and extinction. Behav. Brain Res. 307, 46–53 (2016).
- Intlekofer, K. A. et al. Exercise and sodium butyrate transform a subthreshold learning event into long-term memory via a brain-derived neurotrophic factor-dependent mechanism. Neuropsychopharmacol. Off. Publ. Am. Coll. Neuropsychopharmacol. 38, 2027–2034 (2013).
- Graff, E. C., Fang, H., Wanders, D. & Judd, R. L. Anti-inflammatory effects of the hydroxycarboxylic acid receptor 2. Metabolism. 65, 102–113 (2016).
- Säemann, M. D. et al. Anti-inflammatory effects of sodium butyrate on human monocytes: potent inhibition of IL-12 and up-regulation of IL-10 production. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 14, 2380–2382 (2000).
- Cavaglieri, C. R. et al. Differential effects of short-chain fatty acids on proliferation and production of pro- and anti-inflammatory cytokines by cultured lymphocytes. Life Sci. 73, 1683–1690 (2003).
- Ni, Y.-F. et al. Histone deacetylase inhibitor, butyrate, attenuates lipopolysaccharide-induced acute lung injury in mice. Respir. Res. 11, 33 (2010).
- Usami, M. et al. Butyrate and trichostatin A attenuate nuclear factor kappaB activation and tumor necrosis factor alpha secretion and increase prostaglandin E2 secretion in human peripheral blood mononuclear cells. Nutr. Res. N. Y. N 28, 321–328 (2008).
- Vinolo, M. A. R., Rodrigues, H. G., Nachbar, R. T. & Curi, R. Regulation of Inflammation by Short Chain Fatty Acids. Nutrients 3, 858–876 (2011).
- Arpaia, N. et al. Metabolites produced by commensal bacteria promote peripheral regulatory T-cell generation. Nature 504, 451–455 (2013).
- Smith, P. M. et al. The microbial metabolites, short-chain fatty acids, regulate colonic Treg cell homeostasis. Science 341, 569–573 (2013).
- Furusawa, Y. et al. Commensal microbe-derived butyrate induces the differentiation of colonic regulatory T cells. Nature 504, 446–450 (2013).
- Akimova, T. et al. Histone/protein deacetylase inhibitors increase suppressive functions of human FOXP3+ Tregs. Clin. Immunol. Orlando Fla 136, 348–363 (2010).
- Fontenelle, B. & Gilbert, K. M. n-Butyrate anergized effector CD4+ T cells independent of regulatory T cell generation or activity. Scand. J. Immunol. 76, 457–463 (2012).
- Schauber, J. et al. Histone-deacetylase inhibitors induce the cathelicidin LL-37 in gastrointestinal cells. Mol. Immunol. 41, 847–854 (2004).
- Kida, Y., Shimizu, T. & Kuwano, K. Sodium butyrate up-regulates cathelicidin gene expression via activator protein-1 and histone acetylation at the promoter region in a human lung epithelial cell line, EBC-1. Mol. Immunol. 43, 1972–1981 (2006).
- Schwarz, A., Bruhs, A. & Schwarz, T. The Short-Chain Fatty Acid Sodium Butyrate Functions as a Regulator of the Skin Immune System. J. Invest. Dermatol. 137, 855–864 (2017).
- Karna, E., Trojan, S. & Pałka, J. A. The mechanism of butyrate-induced collagen biosynthesis in cultured fibroblasts. Acta Pol. Pharm. 66, 129–134 (2009).
- Chen, T.-H., Chen, W.-M., Hsu, K.-H., Kuo, C.-D. & Hung, S.-C. Sodium butyrate activates ERK to regulate differentiation of mesenchymal stem cells. Biochem. Biophys. Res. Commun. 355, 913–918 (2007).
- Katono, T. et al. Sodium butyrate stimulates mineralized nodule formation and osteoprotegerin expression by human osteoblasts. Arch. Oral Biol. 53, 903–909 (2008).
- Yan, J. et al. Gut microbiota induce IGF-1 and promote bone formation and growth. Proc. Natl. Acad. Sci. U. S. A. 113, E7554–E7563 (2016).
- Guilloteau, P. et al. From the gut to the peripheral tissues: the multiple effects of butyrate. Nutr. Res. Rev. 23, 366–384 (2010).
- Machiels, K. et al. A decrease of the butyrate-producing species Roseburia hominis and Faecalibacterium prausnitzii defines dysbiosis in patients with ulcerative colitis. Gut 63, 1275–1283 (2014).
- Cao, Y., Shen, J. & Ran, Z. H. Association between Faecalibacterium prausnitzii Reduction and Inflammatory Bowel Disease: A Meta-Analysis and Systematic Review of the Literature. Gastroenterol. Res. Pract. 2014, 872725 (2014).
SCFAs Part 2: The benefits of butyrate
Did you know that butyrate, a metabolite produced in your gut, has benefits for the brain, skin, immune system, and more?
In the last installment of the short-chain fatty acid (SCFA) series, I covered the basics of SCFAs, including how they are produced, where they traffic in the body, and their varied functions. In this second article, I’ll focus on the 4-carbon SCFA butyrate, and discuss the many benefits it has on host health.
Butyrate protects the gut
Butyrate is the preferred substrate for colonocytes, providing 60-70% of the energy requirements for colonic epithelial cells1,2. Butyrate suppresses colonic inflammation,3 is immunoregulatory in the gut,4 and improves gut barrier permeability by accelerating assembly of tight junction proteins.5,6
Butyrate also influences the mucus layer. A healthy colonic epithelium is coated in a double layer of mucus. The thick, inner layer is dense and largely devoid of microbes, protecting the epithelium from contact with commensals and pathogens alike. The loose, outer layer of mucus is home to many bacteria, some of which feed on the glycoproteins of the outer mucus layer itself. Both of these mucus layers are organized by the MUC2 mucin protein, which is secreted by goblet cells in the epithelium. Supplementation of physiological concentrations of butyrate has been shown to increase MUC2 gene expression and MUC2 secretion in a human goblet cell line.7,8
Butyrate improves metabolic function
Butyrate also influences metabolism. In a mouse model of obesity, supplemental butyrate was shown to improve insulin sensitivity, increase energy expenditure, and reduce adiposity. It also increased the number of mitochondria and their activity in skeletal muscle and brown fat tissue.9 This is not surprising, as butyrate responsive receptors are highly expressed in adipose tissue. This expression declines with age, and may explain the age-associated decline in insulin sensitivity.10,11 Butyrate also induces the production of satiety hormones, which reduce food intake.12
Butyrate may also have benefits for other metabolic diseases. In rats, oral butyrate given to young mice early in life showed a trend towards delaying the development of diabetes13. Butyrate supplementation has also been shown to attenuate diet-induced fatty liver disease in mice by improving gut barrier function and correcting microbial dysbiosis14.
Butyrate alters gene expression
While our genetic makeup is essentially unchangeable, the epigenetics, or expression of our genes, is quite malleable. In the nucleus of every cell in our body, DNA is wound around histone proteins. When the histones of a gene are deacetylated, the histones and their associated DNA tend to pack very tightly. This makes it very difficult for any enzymes to bind to transcribe the message encoded in the DNA. The gene will not be made into a protein, and is functionally “off”. On the other hand, when the histones of a gene are acetylated, the histones and their associated DNA tend to pack more loosely. This allows enzymes to bind to the promoter region of the gene and initiate its transcription. This is the first step towards making a functional protein from the gene.
So, what determines if a gene is acetylated or deacetylated? Enzymes called histone acetyltransferases (HATs) add acetyl groups to histones, while enzymes called histone deacetylases (HDACs) remove acetyl groups. Many signaling molecules in the body can influence the activity of these enzymes. One such molecule is…you guessed it…butyrate! Butyrate increases acetylation in two different ways, for distinct sets of genes. First, butyrate is considered a potent HDAC activity inhibitor, meaning that it prevents HDACs from removing acetyl groups (Boffa 1978, Hinnebusch, 2002, Kiefer 2006, Waldecker 2008). For other genes, butyrate enhances HAT activity. Both of these mechanisms serve to increase acetylation and keep genes turned “on”. In the next section, we’ll see how this is put into action in cancer cells.
Butyrate is protective against cancer
Butyrate has long been known to stimulate epithelial cell proliferation, yet at the same time, be protective against colon cancer. In other words, butyrate could exert different effects in healthy and cancerous colonic cells.
Only recently was the mechanism behind this discovered. In an elegant set of experiments, it was determined that the Warburg effect could explain the paradoxical nature of butyrate. The Warburg effect is a phenomenon that involves the metabolism of rapidly-dividing cancerous cells. In a normal epithelial cell, fatty acids like butyrate are largely metabolized via the tricarboxylic acid (TCA) cycle in the mitochondria for energy. An increase in flux through the TCA cycle results in the production of citrate in the cell cytosol, which can be converted to acetyl CoA in the nucleus. This acetyl CoA provides the acetyl group necessary for HATs to acetylate the histones of genes that promote epithelial cell proliferation.
Cancerous cells, on the other hand, tend to have dysfunctional mitochondria, meaning that they can’t metabolize fatty acids (like butyrate) for energy. Because of this, they rely largely on glucose metabolism for their energy production and fatty acids like butyrate tend to accumulate in the cell. This increased cellular butyrate concentration results in increased HDAC inhibition, and the expression of genes that prevents cancer cells from replicating.15
This phenomenon isn’t unique to colon cancer. Butyrate has been identified as having anticancer activity in a variety of human cancer cell lines, including tongue cancer,16 prostate cancer,17,18 liver cancer,19,20 breast cancer,21 lung cancer,22,23 and neuroblastoma.24
Butyrate may benefit the brain
Unfortunately, most of our butyrate-and-the-brain research is limited to animal models. Still, butyrate has been shown to have widespread and dramatic effects on brain health. Mice given sodium butyrate or the butyrate-producing bacteria Clostridium butyricum showed increased neurogenesis, reduced oxidative stress, and improved recovery following ischemic brain injury.25,26 Sodium butyrate has also been shown to restore the integrity of the blood-brain barrier and attenuate neurological deficits following traumatic brain injury in mice.27 Additionally, butyrate may prevent nerve degeneration in the autosomal recessive disease spinal muscular atrophy.28
Butyrate has shown anti-depressant activity in mice, altering expression of genes like brain-derived neurotrophic factor (BDNF) in the hippocampus and frontal cortex and increasing serotonin levels.29,30 It also reduces manic-like behavior in a drug-induced model of bipolar disorder31, and increases expression of neurotrophic factors and reduces cognitive impairment in models of chronic mild stress and maternal separation.32
Butyrate also has interesting implications for memory formation and retention. Disturbances in hippocampal gastrin-releasing peptide receptors (GRPRs) may inhibit proper memory formation and extinction and contribute to the cognitive impairment associated with neurodevelopmental disorders. One study in mice found that HDAC inhibition by butyrate could attenuate the cognitive impairment that results from GRPR signaling blockade.33 Another study found that, like exercise, butyrate places the brain in a “state of readiness for plasticity”.34
Lastly, butyrate may have benefits for autism spectrum disorder. I’ll be discussing this more in part 5 when I discuss SCFA imbalance and the propionate-ASD connection.
Butyrate regulates the immune system
Butyrate also plays a major role in regulating the immune system, and butyrate responsive receptors have been found on virtually every type of immune cell.35
Butyrate signaling results in an overall anti-inflammatory immune profile, inhibiting production of proinflammatory cytokine and upregulating production of anti-inflammatory cytokines36,37. It has been shown to do this even in the face of an inflammatory insult, such as lipopolysaccharide (LPS) from the cell walls of gram-negative bacteria. This may occur through HDAC inhibition of the NF-kB pathway, a major pathway involved in the release of inflammatory cytokines38,39. In fact, butyrate mediated HDAC inhibition has been observed in many different immune cell types.40
Butyrate also promotes the generation of colonic and peripheral regulatory T cells (Tregs), which serve to help dampen the immune response,41–43 influences both Treg and effector T cell function,44,45 and induces activity of cathelicidin, a potent antimicrobial known to play an important role in innate defense against bacterial infection, in colonic and lung epithelial cells.46,47
Butyrate and the skin
Of particular interest to me, due to my previous history with eczema and laboratory research on the gut-skin axis, is the effect of butyrate on skin health. Butyrate acts as a regulator of the skin immune system. Subcutaneously or topically applied butyrate was shown to reduce contact hypersensitivity, likely explained by the observed increase in regulatory T cells and reduction in inflammatory effector T cells.48 Butyrate also induces the synthesis of collagen, an important structural component of skin.49
Butyrate and bone health
Butyrate may also improve bone health. Bone is constantly being remodeled in a delicate balance between osteoblast (bone-building) cell activity and osteoclast (bone breakdown) cell activity. Mesenchymal stem cells (MSCs), which reside in the bone marrow, can differentiate into osteoblasts (bone-building cells) or adipocytes (fat cells). One study found that sodium butyrate caused MSCs to preferentially differentiate into osteoblasts, suggesting that it could promote bone formation.50 Another study found that osteoblasts cultured with butyrate had increased number and calcium content of mineralized nodules. They also expressed higher levels of osteoprotegerin, a protein that serves to reduce osteoclast (bone breakdown) activity. 51 In animal models, a mixture of SCFAs that includes butyrate has been shown to increase circulating levels of insulin-like growth factor (IGF) and promote bone formation.52
Ways to increase butyrate
To sum up, we’ve learned that butyrate is a potent modifier of gene expression, and has widespread benefits for host health. There are many reasons we might want to increase butyrate production, and there are several ways to do so:
- Increase the abundance of butyrate-producing bacteria. Bacterial species with particularly high capacities for butyrate production belong to both the Firmicutes and Bacteroidetes phyla and include species such as Clostridium leptum, Eubacterium spp., Roseburia spp., Coprococcus spp., and Faecalibacterium prausnitzii.53 Genera like Bifidobacteria and Lactobaccilus may also contribute to butyrate production by producing acetate and lactate, which other bacteria can then convert to butyrate.
- Increase the substrates available to butyrate-producing bacteria. This means eating prebiotic foods with fermentable fibers that stimulate butyrate production. Some of the most potently butyrogenic fibers include resistant starch and fructooligosaccharides. I’ll be discussing this more in SCFAs part 7: the definitive guide to fiber.
- Supplementation. This should be done with extreme caution, and with the right supplement form. Which brings me to my final point for today…
Is butyrate always good?
Nope! This is only part 2! In the next article of this series, I’ll talk about how excess butyrate can be toxic, particularly in conditions involving gut inflammation. I’ll also discuss the various forms of supplementation, and why most butyrate supplements may do more harm than good. Be sure to subscribe below!
Sources:
- Donohoe, D. R. et al. The Microbiome and Butyrate Regulate Energy Metabolism and Autophagy in the Mammalian Colon. Cell Metab. 13, 517–526 (2011).
- Suzuki, T., Yoshida, S. & Hara, H. Physiological concentrations of short-chain fatty acids immediately suppress colonic epithelial permeability. Br. J. Nutr. 100, 297–305 (2008).
- Singh, N. et al. Activation of Gpr109a, receptor for niacin and the commensal metabolite butyrate, suppresses colonic inflammation and carcinogenesis. Immunity 40, 128–139 (2014).
- Chang, P. V., Hao, L., Offermanns, S. & Medzhitov, R. The microbial metabolite butyrate regulates intestinal macrophage function via histone deacetylase inhibition. Proc. Natl. Acad. Sci. U. S. A. 111, 2247–2252 (2014).
- Peng, L., He, Z., Chen, W., Holzman, I. R. & Lin, J. Effects of butyrate on intestinal barrier function in a Caco-2 cell monolayer model of intestinal barrier. Pediatr. Res. 61, 37–41 (2007).
- Peng, L., Li, Z.-R., Green, R. S., Holzman, I. R. & Lin, J. Butyrate Enhances the Intestinal Barrier by Facilitating Tight Junction Assembly via Activation of AMP-Activated Protein Kinase in Caco-2 Cell Monolayers. J. Nutr. 139, 1619–1625 (2009).
- Burger-van Paassen, N. et al. The regulation of intestinal mucin MUC2 expression by short-chain fatty acids: implications for epithelial protection. Biochem. J. 420, 211–219 (2009).
- Hatayama, H., Iwashita, J., Kuwajima, A. & Abe, T. The short chain fatty acid, butyrate, stimulates MUC2 mucin production in the human colon cancer cell line, LS174T. Biochem. Biophys. Res. Commun. 356, 599–603 (2007).
- Gao, Z. et al. Butyrate improves insulin sensitivity and increases energy expenditure in mice. Diabetes 58, 1509–1517 (2009).
- Thangaraju, M. et al. GPR109A is a G-protein-coupled receptor for the bacterial fermentation product butyrate and functions as a tumor suppressor in colon. Cancer Res. 69, 2826–2832 (2009).
- Fink, R. I., Kolterman, O. G., Griffin, J. & Olefsky, J. M. Mechanisms of Insulin Resistance in Aging. J. Clin. Invest. 71, 1523–1535 (1983).
- Lin, H. V. et al. Butyrate and Propionate Protect against Diet-Induced Obesity and Regulate Gut Hormones via Free Fatty Acid Receptor 3-Independent Mechanisms. PLOS ONE 7, e35240 (2012).
- Li, N. et al. Butyrate and type 1 diabetes mellitus: can we fix the intestinal leak? J. Pediatr. Gastroenterol. Nutr. 51, 414–417 (2010).
- Zhou, D. et al. Sodium butyrate attenuates high-fat diet-induced steatohepatitis in mice by improving gut microbiota and gastrointestinal barrier. World J. Gastroenterol. 23, 60–75 (2017).
- Donohoe, D. R. et al. The Warburg effect dictates the mechanism of butyrate-mediated histone acetylation and cell proliferation. Mol. Cell 48, 612–626 (2012).
- Jeng, J.-H. et al. Toxic and metabolic effect of sodium butyrate on SAS tongue cancer cells: role of cell cycle deregulation and redox changes. Toxicology 223, 235–247 (2006).
- Kim, J., Park, H., Im, J. Y., Choi, W. S. & Kim, H. S. Sodium butyrate regulates androgen receptor expression and cell cycle arrest in human prostate cancer cells. Anticancer Res. 27, 3285–3292 (2007).
- Kuefer, R. et al. Sodium butyrate and tributyrin induce in vivo growth inhibition and apoptosis in human prostate cancer. Br. J. Cancer 90, 535–541 (2004).
- Nakamura, M. et al. Reduction of telomerase activity in human liver cancer cells by a histone deacetylase inhibitor. J. Cell. Physiol. 187, 392–401 (2001).
- Wakabayashi, K. et al. Gene expression associated with the decrease in malignant phenotype of human liver cancer cells following stimulation with a histone deacetylase inhibitor. Int. J. Oncol. 26, 233–239 (2005).
- M, M. & R, K. Bcl-2 expression regulates sodium butyrate-induced apoptosis in human MCF-7 breast cancer cells. Cell Growth Differ. Mol. Biol. J. Am. Assoc. Cancer Res. 7, 311–318 (1996).
- Pellizzaro, C., Coradini, D., Daniotti, A., Abolafio, G. & Daidone, M. G. Modulation of cell cycle-related protein expression by sodium butyrate in human non-small cell lung cancer cell lines. Int. J. Cancer 91, 654–657 (2001).
- Denlinger, C. E., Keller, M. D., Mayo, M. W., Broad, R. M. & Jones, D. R. Combined proteasome and histone deacetylase inhibition in non–small cell lung cancer. J. Thorac. Cardiovasc. Surg. 127, 1078–1086 (2004).
- Schneider, F. H. Effects of sodium butyrate on mouse neuroblastoma cells in culture. Biochem. Pharmacol. 25, 2309–2317 (1976).
- Kim, H. J., Leeds, P. & Chuang, D.-M. The HDAC inhibitor, sodium butyrate, stimulates neurogenesis in the ischemic brain. J. Neurochem. 110, 1226–1240 (2009).
- Sun, J. et al. Clostridium butyricum pretreatment attenuates cerebral ischemia/reperfusion injury in mice via anti-oxidation and anti-apoptosis. Neurosci. Lett. 613, 30–35 (2016).
- Li, H. et al. Sodium butyrate exerts neuroprotective effects by restoring the blood-brain barrier in traumatic brain injury mice. Brain Res. 1642, 70–78 (2016).
- Chang, J.-G. et al. Treatment of spinal muscular atrophy by sodium butyrate. Proc. Natl. Acad. Sci. U. S. A. 98, 9808–9813 (2001).
- Schroeder, F. A., Lin, C. L., Crusio, W. E. & Akbarian, S. Antidepressant-like effects of the histone deacetylase inhibitor, sodium butyrate, in the mouse. Biol. Psychiatry 62, 55–64 (2007).
- Sun, J. et al. Antidepressant-like effects of sodium butyrate and its possible mechanisms of action in mice exposed to chronic unpredictable mild stress. Neurosci. Lett. 618, 159–166 (2016).
- Varela, R. B. et al. Sodium butyrate and mood stabilizers block ouabain-induced hyperlocomotion and increase BDNF, NGF and GDNF levels in brain of Wistar rats. J. Psychiatr. Res. 61, 114–121 (2015).
- Valvassori, S. S. et al. Sodium butyrate functions as an antidepressant and improves cognition with enhanced neurotrophic expression in models of maternal deprivation and chronic mild stress. Curr. Neurovasc. Res. 11, 359–366 (2014).
- Petry, F. S. et al. Histone deacetylase inhibition prevents the impairing effects of hippocampal gastrin-releasing peptide receptor antagonism on memory consolidation and extinction. Behav. Brain Res. 307, 46–53 (2016).
- Intlekofer, K. A. et al. Exercise and sodium butyrate transform a subthreshold learning event into long-term memory via a brain-derived neurotrophic factor-dependent mechanism. Neuropsychopharmacol. Off. Publ. Am. Coll. Neuropsychopharmacol. 38, 2027–2034 (2013).
- Graff, E. C., Fang, H., Wanders, D. & Judd, R. L. Anti-inflammatory effects of the hydroxycarboxylic acid receptor 2. Metabolism. 65, 102–113 (2016).
- Säemann, M. D. et al. Anti-inflammatory effects of sodium butyrate on human monocytes: potent inhibition of IL-12 and up-regulation of IL-10 production. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 14, 2380–2382 (2000).
- Cavaglieri, C. R. et al. Differential effects of short-chain fatty acids on proliferation and production of pro- and anti-inflammatory cytokines by cultured lymphocytes. Life Sci. 73, 1683–1690 (2003).
- Ni, Y.-F. et al. Histone deacetylase inhibitor, butyrate, attenuates lipopolysaccharide-induced acute lung injury in mice. Respir. Res. 11, 33 (2010).
- Usami, M. et al. Butyrate and trichostatin A attenuate nuclear factor kappaB activation and tumor necrosis factor alpha secretion and increase prostaglandin E2 secretion in human peripheral blood mononuclear cells. Nutr. Res. N. Y. N 28, 321–328 (2008).
- Vinolo, M. A. R., Rodrigues, H. G., Nachbar, R. T. & Curi, R. Regulation of Inflammation by Short Chain Fatty Acids. Nutrients 3, 858–876 (2011).
- Arpaia, N. et al. Metabolites produced by commensal bacteria promote peripheral regulatory T-cell generation. Nature 504, 451–455 (2013).
- Smith, P. M. et al. The microbial metabolites, short-chain fatty acids, regulate colonic Treg cell homeostasis. Science 341, 569–573 (2013).
- Furusawa, Y. et al. Commensal microbe-derived butyrate induces the differentiation of colonic regulatory T cells. Nature 504, 446–450 (2013).
- Akimova, T. et al. Histone/protein deacetylase inhibitors increase suppressive functions of human FOXP3+ Tregs. Clin. Immunol. Orlando Fla 136, 348–363 (2010).
- Fontenelle, B. & Gilbert, K. M. n-Butyrate anergized effector CD4+ T cells independent of regulatory T cell generation or activity. Scand. J. Immunol. 76, 457–463 (2012).
- Schauber, J. et al. Histone-deacetylase inhibitors induce the cathelicidin LL-37 in gastrointestinal cells. Mol. Immunol. 41, 847–854 (2004).
- Kida, Y., Shimizu, T. & Kuwano, K. Sodium butyrate up-regulates cathelicidin gene expression via activator protein-1 and histone acetylation at the promoter region in a human lung epithelial cell line, EBC-1. Mol. Immunol. 43, 1972–1981 (2006).
- Schwarz, A., Bruhs, A. & Schwarz, T. The Short-Chain Fatty Acid Sodium Butyrate Functions as a Regulator of the Skin Immune System. J. Invest. Dermatol. 137, 855–864 (2017).
- Karna, E., Trojan, S. & Pałka, J. A. The mechanism of butyrate-induced collagen biosynthesis in cultured fibroblasts. Acta Pol. Pharm. 66, 129–134 (2009).
- Chen, T.-H., Chen, W.-M., Hsu, K.-H., Kuo, C.-D. & Hung, S.-C. Sodium butyrate activates ERK to regulate differentiation of mesenchymal stem cells. Biochem. Biophys. Res. Commun. 355, 913–918 (2007).
- Katono, T. et al. Sodium butyrate stimulates mineralized nodule formation and osteoprotegerin expression by human osteoblasts. Arch. Oral Biol. 53, 903–909 (2008).
- Yan, J. et al. Gut microbiota induce IGF-1 and promote bone formation and growth. Proc. Natl. Acad. Sci. U. S. A. 113, E7554–E7563 (2016).
- Guilloteau, P. et al. From the gut to the peripheral tissues: the multiple effects of butyrate. Nutr. Res. Rev. 23, 366–384 (2010).
- Machiels, K. et al. A decrease of the butyrate-producing species Roseburia hominis and Faecalibacterium prausnitzii defines dysbiosis in patients with ulcerative colitis. Gut 63, 1275–1283 (2014).
- Cao, Y., Shen, J. & Ran, Z. H. Association between Faecalibacterium prausnitzii Reduction and Inflammatory Bowel Disease: A Meta-Analysis and Systematic Review of the Literature. Gastroenterol. Res. Pract. 2014, 872725 (2014).
I am confused about butyrate and acetyl groups. If one has excessive acetaldehyde, is butyrate a good or bad thing to supplement?
Dónde y en que producto conseguir todas estas cepas productoras de butirato…. cada microbioma es diferente y las necesidades son diferentes, que dificil y lamentable, muchos niños y jóvenes con necesidad urgente…
Sodium butyrate orally and by enema really helped get my son back on his feet. It wasn’t sustainable long-term, but it could help many in a severe flare. Now that my son has an ileostomy, I have tried to think through how to best heal the colon when it is cut off from the digestive system so we can attempt a reversal. Do you ever get into that topic?
Thanks for your comment, Lynn. Unfortunately, there is not much literature on colon nutritional support for ileostomy reversal. I would think butyrate enemas might be helpful in this case as well to support colonic epithelial health, but there is no available research to go on in this regard.
Hi Lucy, I have been looking for part 7 of this series as you mentioned it: “I’ll be discussing this more in SCFAs part 7: the definitive guide to fiber.”
Is there a link I’m not able to see? Can’t wait to read!
Hi Paula – so glad you are enjoying the series! I never did get around to part 6 or 7 but hope to write them soon!
Awe, Lucy! I would love to help ya if you need a hand ;) But I reaaaally, really would love to see those 2 more parts posted soon. ;))))
[…] shahii, Roseburia intestinalis, and Coprococcus spp. Notably, these are all microbes that produce butyrate, an essential metabolite for the health of the gut […]
[…] shahii, Roseburia intestinalis, and Coprococcus spp. Notably, these are all microbes that produce butyrate, an essential metabolite for the health of the gut […]